CryoEM structures of AL amyloid fibrils
Two manuscripts were published last week describing high-resolution 3D structures of light chain amyloid fibrils extracted from patient heart tissue. Swuec and coworkers solved the structure of a fibril derived from a lambda 6-57 light chain using cryo electron microscopy (cryoEM); Radameker and coworkers did the same for a lambda 1-44 light chain. These structures provide important information about how amyloid forms in patients. However, the structures themselves are remarkably different, which shows just how much more we have to learn.
Amyloid fibrils are large and non-crystalline, which makes them difficult to study using the structural techniques developed for soluble proteins. So although there have been some important insights, the detailed molecular structures of amyloid fibrils from diseased tissue have remained out of reach. That changed in 2017 with the publication of structures of tau filaments from the brain of a patient who had died of Alzheimer’s disease. A series of technological advances in EM hardware and software over the last few years has enabled this work. It’s great to see it applied to AL amyloidosis. If you’re interested in the really diving into the details of cryoEM, Caltech has a free, detailed online course.
Both fibrils were extracted from heart tissue using repeated washes with water and buffer to remove non-amyloid material. Amyloid is pretty tough and survives multiple rounds of homogenization in a blender. Isolated fibrils were suspended in an ultrathin film of water, then rapidly frozen in liquid ethane. The fast freezing prevents ice crystals forming, which would damage the fibrils.
Structure determination by EM relies on the repetitive nature of the fibrils. Gigabytes of EM images are analyzed to identify individual fibrils, which are then sliced into segments. Each segment is a projection of the fibril, rotated according to its helical twist. By combining many, many segment images, it’s possible to reconstruct the 3D structure that created the projection. Ten years ago, the best reconstructions were intricate blobs that showed where the protein was, but not which bit of protein was where. The new generation of structures can identify specific sidechains and their interactions.
Both fibrils consist of a single protofilament, formed from residues of the variable domain. As has been observed in other fibrils, the constant domain has been partly removed, presumably by proteolysis. Parts of both protein chains are missing from the resolved density, which probably means that they are less well-structured than the resolved core of the fibrils. Both structures have a complex, non-native arrangement of β-strands that makes up the core of the fibril. The native disulfide bond is present in the core of the structures, but the rest of the protein chains are arranged into a series of parallel, in-register cross-beta structures, where the hydrogen bonds run along the long fibril axis. Here are the lambda 1-44 fibril and the lambda 6-57 fibril depicted as topology diagrams.
This non-native structure indicates that the V-domain must unfold in order to form aggregates, consistent with a role for reduced LC stability. This is highlighted in the lambda 1-44 fibril because the orientation of the beta strands around the disulfide are reversed relative to their orientation in the native state of the fibril. Importantly, the complementarity-determining regions of the V-domain pack into a complex structure within the fibril core. This indicates that different LC sequences will alter the stability of the fibril core, which may also constrain which patient LC sequences are able to form amyloid in vivo.
Beyond these similarities though, the structures are remarkably distinct. The regions of beta structure and the overall packing of the cross-beta elements are different. One N-terminus is buried right in the center of the fibril, while the other is unstructured. (This highlights the importance of using native-like termini in biochemical studies – would a his-tagged version of this protein form the same structures?) The lambda 6-57 fibril does not have resolved density for residues 37-65, but these are structured in the lambda 1-44 fibril.
To get a better sense of how different the structures are, I aligned the sequences of the two light chains and looked at where the beta strands are in the native proteins and the fibrils. (See the figure – I took the data from the figures in the papers.) The sequences are 63% identical and the native beta structures are essentially the same as each other. But the structured regions of the fibril and the patterns of beta strands – the arrows above the sequences – are very different.
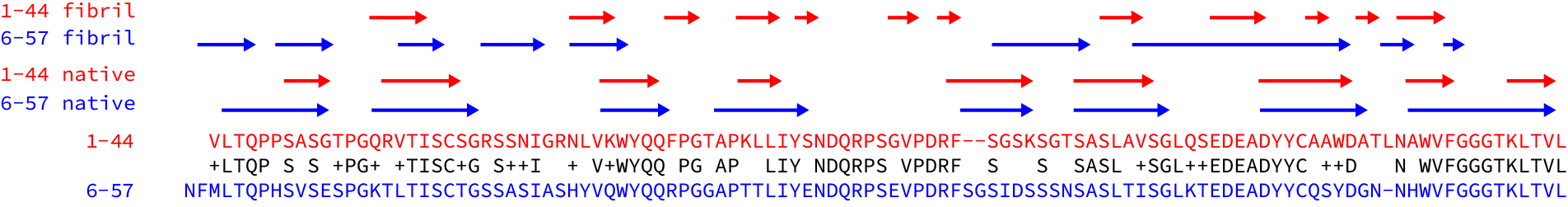
These differences raise a lot of questions. Will every amyloid fibril be unique? Can the same light chain form different structures? Are fibrils in different tissues more, or less similar than fibrils from different patients? Is the structure of a fibril determined primarily by its sequence, or are interactions with other molecules important? Are kappa fibrils different to lambda fibrils?
What’s also interesting is what the structures don’t show. There’s almost no trace of native-like structure in either fibril, which means that the native light chain probably has to unfold completely in order to aggregate. This rules out a large class of models for fibrils that retained some of the native structure. In addition, there’s nothing that would obviously cause toxicity. And there’s very little in common between the fibrils that could explain why they cause organ dysfunction in similar ways.
As ever, there’s plenty more work to be done. But these structures represent an important milestone. It’s really helpful to have a detailed model of what a structure might look like in order to ask what different parts of it might be doing. Congratulations to both sets of authors!